Part 2: l'essaim, un cerveau virtuel?
On compare souvent les sociétés d’insectes à des “super-organismes” tant leur fonctionnement semble doté d’une vie propre, grâce à l’extraordinaire coordination des bestioles qui les composent. Dans son livre "Gödel Escher Bach", Douglas Hofstadter va un cran plus loin lorsqu'il évoque une fourmilière (Mme de Montfourmi) avec laquelle le fourmilier passe de longues soirées à bavarder, alors que chaque fourmi qui la compose est parfaitement stupide et a même une peur terrible de se faire manger par le fourmilier. "Il me semble, écrit-il, que la situation présente des analogies avec la composition d'un cerveau humain, avec ses neurones. Personne n'oserait soutenir que chacune des cellules du cerveau doit être une entité intelligente pour expliquer qu'une personne puisse avoir une conversation intelligente" (p353).
Si l’on en juge le mécanisme de décision des abeilles en quête d’un nouveau nid dont je vous ai parlé la dernière fois, Hofstadter pourrait avoir vu juste. L’intelligence qui s’en dégage (mesurable à la pertinence des choix) est incomparablement supérieure à celle que pourrait produire une abeille prise individuellement : en deux jours, l’essaim arrive à dégotter le meilleur site à des kilomètres à la ronde puis à se mettre d’accord pour y emménager. Se pourrait-il que l’essaim partage des points commun avec le mode de fonctionnement d’un cerveau de vertébré? C’est l’idée que défend Thomas Seeley.
Comment on choisit dans sa tête
Prenez un singe de bonne composition et placez-le devant un écran sur lesquel défilent plein de petites flèches pointant dans toutes les directions de façon aléatoire (↗←↑↖→↙↓↘) . Entraînez votre singe à détecter si durant une séquence elles pointent plutôt vers la droite ou plutôt vers la gauche et de l’indiquer par un mouvement des yeux dans cette direction.
Les chercheurs en neurologie ont décomposé ce qui se passe dans le cerveau au cours de la séquence:
1) Une première catégorie de neurones (dans le lobe temporal médian - MT) focalisés sur une aire particulière du champ visuel, détectent la direction des flèches dans cette zone. Certains déchargent quand ils détectent une flèche vers la droite (↗ ↘ →), d’autres quand la flèche pointe vers la gauche (← ↖ ↙)
2) Cette information est transmise à une seconde catégorie de neurones (dans le lobe latéral intrapariétal LIP), également organisés par direction (droite ou gauche) et qui intègrent la quantité de décharges reçus par les neurones MT correspondant à cette direction. Plus les neurones LIP reçoivent de stimulation de la part des neurones MT codant pour la même direction qu’eux, plus ils déchargent. Et plus ils déchargent plus ils inhibent les neurones LIP codant pour l’autre direction, si bien qu’au bout d’un moment, s’il y a par exemple plus de flèches pointant vers la gauche, seuls les neurones LIP codant pour la gauche déchargent comme des fous. Le mécanisme d’inhibition entre neurones LIP fait ainsi bien ressortir le “contraste” entre les proportions.
3) Lorsque le taux de décharge d’une catégorie de neurones LIP dépasse un certain seuil, ils déclenchent l’activation des neurones moteurs qui pilotent le mouvement des yeux (vers la gauche en l’occurrence). Et voilà!
On retrouve à peu près les mêmes mécanismes que dans notre essaim d’abeilles!
- les neurones MT, détecteurs de direction dans une zone particulière, sont l’équivalent des abeilles exploratrices qui signalent par leur danse la valeur d’un nouveau site dans une région donnée
- les neurones LIP, qui intègrent les signaux MT sont équivalents au nombre d’abeilles qui visitent un site donné, puisque ce nombre reflète finalement la quantité de publicité cumulée dans les dernières heures pour ce site. Les deux types d’intégrateurs ont la particularité d’être “poreux”, de perdre de l’information à mesure que le temps passe. Pour les abeilles, cette porosité prend la forme d’une “lassitude” graduelle pour un site déjà visité plusieurs fois et elle joue un rôle clé pour le choix puisque c’est grâce à elle que s’éliminent les seconds choix et qu’un consensus émerge rapidement. Plus de “porosité” permet d’arriver plus rapidement à une décision avec le risque de rater l’optimum. Moins de porosité améliore la qualité de la décision mais est plus coûteux en temps. Bref, cette porosité modélise l’impulsivité de la décision.
- Enfin dans les deux cas, la décision se prend lorsqu’un des intégrateurs dépasse un seuil critique d’activation. Ce seuil est lui aussi un paramètre important puisqu’il influe sur la rapidité avec laquelle la décision est prise.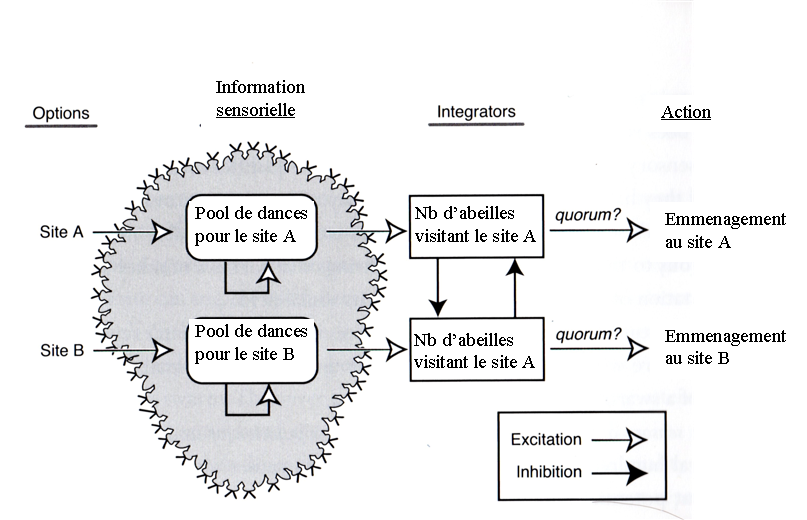
Il y a juste un détail sur lequel on reste sur sa faim, c’est la question de l’inhibition mutuelle chez les abeilles entre les différents choix. Dans le cerveau les neurones LIP s’inhibent mutuellement à proportion de leur excitation respective alors qu’on a vu qu’il n’y a pas d’inhibition mutuelle entre les différents choix des abeilles. Thomas Seeley suggère qu’une telle inhibition existe dans la mesure où il n’y a qu’un pool limité d’abeilles hésitantes. Plus il y a d’abeilles recrutées pour un excellent site, moins il en reste de recrutables pour un autre.
Même si je ne suis pas hyper convaincu par l’argument, la démonstration de Seeley reste globalement convaincante qu’il existe de grandes similitudes entre ces deux machines à prendre des décisions que sont le cerveau et un essaim d’abeilles. Il me semble qu’on a là un modèle stochastique particulièrement prometteur pour développer des systèmes d’aide à la décision. Au point que je me suis longtemps demandé pourquoi les sociétés d’abeilles n’avaient pas collectivement évolué encore plus loin, se transformant en êtres supérieurement intelligents, capables de faire collectivement bien mieux que nous, en termes de déplacement (ça vole), de défense (plus flexible que Mask!). Avec l’immortalité en plus, remarquez bien, car l’essaim se régénère en permanence, remplaçant ses blessures par des insectes tout neufs (y compris la reine, on peut supposer qu’il y en aurait toujours une de rechange au cas où). Bref pourquoi les sociétés d’abeilles, de fourmis et de termites n’ont-elles pas progressé plus vite que nous, du point de vue de l’intelligence, de la créativité, de la capacité d’apprentissage etc? Et pourquoi restent-ils des guêpes non sociales limitées par la taille de leur cerveau minuscule?
Je pense avoir trouvé la réponse: c’est que tous ces insectes sociaux n’ont pas inventé l’électricité. Je m’explique: la transmission de l’information entre insectes par des phéromones, des alertes visuelles ou des contacts est certes comparable au principe d’un influx nerveux. Mais ces signaux chimiques ou comportementaux se propagent infiniment moins vite qu’un signal électrique transitant d’un neurone à l’autre! Il faut quelques secondes à une abeille pour informer ses collègues sur la qualité d’un site, sa distance et la direction où il se trouve, alors que ça ne prend que quelques millisecondes à nos neurones. Avec un cerveau fonctionnant sans électricité, on ne va pas bien loin! Si d’aventure un essaim d’insectes sociaux mutants se prenait pour Hulk, le monstre en question serait un peu lent à la détente et ça ferait un très mauvais film d’horreur. Je ne sais pas ce que vaut cette explication, mais au moins ça me permettra de dormir tranquille...
Si l’on en juge le mécanisme de décision des abeilles en quête d’un nouveau nid dont je vous ai parlé la dernière fois, Hofstadter pourrait avoir vu juste. L’intelligence qui s’en dégage (mesurable à la pertinence des choix) est incomparablement supérieure à celle que pourrait produire une abeille prise individuellement : en deux jours, l’essaim arrive à dégotter le meilleur site à des kilomètres à la ronde puis à se mettre d’accord pour y emménager. Se pourrait-il que l’essaim partage des points commun avec le mode de fonctionnement d’un cerveau de vertébré? C’est l’idée que défend Thomas Seeley.
Comment on choisit dans sa tête
Prenez un singe de bonne composition et placez-le devant un écran sur lesquel défilent plein de petites flèches pointant dans toutes les directions de façon aléatoire (↗←↑↖→↙↓↘) . Entraînez votre singe à détecter si durant une séquence elles pointent plutôt vers la droite ou plutôt vers la gauche et de l’indiquer par un mouvement des yeux dans cette direction.
Les chercheurs en neurologie ont décomposé ce qui se passe dans le cerveau au cours de la séquence:
1) Une première catégorie de neurones (dans le lobe temporal médian - MT) focalisés sur une aire particulière du champ visuel, détectent la direction des flèches dans cette zone. Certains déchargent quand ils détectent une flèche vers la droite (↗ ↘ →), d’autres quand la flèche pointe vers la gauche (← ↖ ↙)
2) Cette information est transmise à une seconde catégorie de neurones (dans le lobe latéral intrapariétal LIP), également organisés par direction (droite ou gauche) et qui intègrent la quantité de décharges reçus par les neurones MT correspondant à cette direction. Plus les neurones LIP reçoivent de stimulation de la part des neurones MT codant pour la même direction qu’eux, plus ils déchargent. Et plus ils déchargent plus ils inhibent les neurones LIP codant pour l’autre direction, si bien qu’au bout d’un moment, s’il y a par exemple plus de flèches pointant vers la gauche, seuls les neurones LIP codant pour la gauche déchargent comme des fous. Le mécanisme d’inhibition entre neurones LIP fait ainsi bien ressortir le “contraste” entre les proportions.
3) Lorsque le taux de décharge d’une catégorie de neurones LIP dépasse un certain seuil, ils déclenchent l’activation des neurones moteurs qui pilotent le mouvement des yeux (vers la gauche en l’occurrence). Et voilà!
On retrouve à peu près les mêmes mécanismes que dans notre essaim d’abeilles!
- les neurones MT, détecteurs de direction dans une zone particulière, sont l’équivalent des abeilles exploratrices qui signalent par leur danse la valeur d’un nouveau site dans une région donnée
- les neurones LIP, qui intègrent les signaux MT sont équivalents au nombre d’abeilles qui visitent un site donné, puisque ce nombre reflète finalement la quantité de publicité cumulée dans les dernières heures pour ce site. Les deux types d’intégrateurs ont la particularité d’être “poreux”, de perdre de l’information à mesure que le temps passe. Pour les abeilles, cette porosité prend la forme d’une “lassitude” graduelle pour un site déjà visité plusieurs fois et elle joue un rôle clé pour le choix puisque c’est grâce à elle que s’éliminent les seconds choix et qu’un consensus émerge rapidement. Plus de “porosité” permet d’arriver plus rapidement à une décision avec le risque de rater l’optimum. Moins de porosité améliore la qualité de la décision mais est plus coûteux en temps. Bref, cette porosité modélise l’impulsivité de la décision.
- Enfin dans les deux cas, la décision se prend lorsqu’un des intégrateurs dépasse un seuil critique d’activation. Ce seuil est lui aussi un paramètre important puisqu’il influe sur la rapidité avec laquelle la décision est prise.
Il y a juste un détail sur lequel on reste sur sa faim, c’est la question de l’inhibition mutuelle chez les abeilles entre les différents choix. Dans le cerveau les neurones LIP s’inhibent mutuellement à proportion de leur excitation respective alors qu’on a vu qu’il n’y a pas d’inhibition mutuelle entre les différents choix des abeilles. Thomas Seeley suggère qu’une telle inhibition existe dans la mesure où il n’y a qu’un pool limité d’abeilles hésitantes. Plus il y a d’abeilles recrutées pour un excellent site, moins il en reste de recrutables pour un autre.
Même si je ne suis pas hyper convaincu par l’argument, la démonstration de Seeley reste globalement convaincante qu’il existe de grandes similitudes entre ces deux machines à prendre des décisions que sont le cerveau et un essaim d’abeilles. Il me semble qu’on a là un modèle stochastique particulièrement prometteur pour développer des systèmes d’aide à la décision. Au point que je me suis longtemps demandé pourquoi les sociétés d’abeilles n’avaient pas collectivement évolué encore plus loin, se transformant en êtres supérieurement intelligents, capables de faire collectivement bien mieux que nous, en termes de déplacement (ça vole), de défense (plus flexible que Mask!). Avec l’immortalité en plus, remarquez bien, car l’essaim se régénère en permanence, remplaçant ses blessures par des insectes tout neufs (y compris la reine, on peut supposer qu’il y en aurait toujours une de rechange au cas où). Bref pourquoi les sociétés d’abeilles, de fourmis et de termites n’ont-elles pas progressé plus vite que nous, du point de vue de l’intelligence, de la créativité, de la capacité d’apprentissage etc? Et pourquoi restent-ils des guêpes non sociales limitées par la taille de leur cerveau minuscule?
Je pense avoir trouvé la réponse: c’est que tous ces insectes sociaux n’ont pas inventé l’électricité. Je m’explique: la transmission de l’information entre insectes par des phéromones, des alertes visuelles ou des contacts est certes comparable au principe d’un influx nerveux. Mais ces signaux chimiques ou comportementaux se propagent infiniment moins vite qu’un signal électrique transitant d’un neurone à l’autre! Il faut quelques secondes à une abeille pour informer ses collègues sur la qualité d’un site, sa distance et la direction où il se trouve, alors que ça ne prend que quelques millisecondes à nos neurones. Avec un cerveau fonctionnant sans électricité, on ne va pas bien loin! Si d’aventure un essaim d’insectes sociaux mutants se prenait pour Hulk, le monstre en question serait un peu lent à la détente et ça ferait un très mauvais film d’horreur. Je ne sais pas ce que vaut cette explication, mais au moins ça me permettra de dormir tranquille...
Source: Thomas Seeley, Honeybee Democracy
La première partie de la saga ici.