On a vu dans le dernier billet pourquoi l'image de gènes programmant mécaniquement les organismes qui les hébergent est loin de la réalité. L'ADN n'est pas un "programmateur" mais un code passif, que la machinerie cellulaire peut lire de plusieurs manière selon l'environnement chimique de la cellule. Du coup, un même code génétique peut produire des formes morphologiques très altérées en cas de forte perturbation du milieu.
Cette combinaison de robustesse et de plasticité en cas de gros pépin pourrait presque être vue comme une caractéristique du vivant, au même titre que la capacité à se reproduire ou à se développer...
L'assimilation génétique
Si cette idée n'a rien d'évident, c'est que la sensibilité des organismes à l'environnement ne saute pas vraiment aux yeux. Les organismes d'une même espèce se ressemblent plutôt beaucoup et ils ne se transforment pas en mutants garous dès qu'il pleut ou qu'il fait chaud. Dans les années 1940, le biologiste anglais Conrad Waddington eut l’idée que cette relative immuabilité était en réalité due à l’action efficace de mécanismes régulateurs (homéostatiques, comme on dit) qui maintiennent constant le milieu intérieur de la cellule et qui corrigent ou atténuent les erreurs de lecture des gènes.
Il avait remarqué qu’il suffisait de soumettre des larves de moucherons (toujours la même bonne drosophila melanogaster) à des chocs thermiques pendant quelques minutes pour qu’ils présentent, une fois adultes, des anomalies dans les nervures des ailes. Anomalies qu'on observe très rarement en temps normal (0,5% des cas). En sélectionnant les individus anormaux et en répétant la même manip sur plusieurs générations, Waddington parvint à obtenir une lignée de moucherons présentant toutes ces anomalies, même en l'absence de choc thermique! Hérédité des caractère acquis pensez-vous? Pas forcément.
 |
| L'anomalie dans les nervures de l'aile d'une drosophile adulte (source ici) |
“The constancy of the wild-type must be taken as evidence of the buffering of the genotype against minor variations, not only in the environment in which the animals developped, but also in its genetic make-up”.
Ce qu’on pourrait traduire par “L’immuabilité des espèces à l’état sauvage montre que le génotype est immunisé contre les petites variations, celles du milieu dans lequel se développent les animaux, mais aussi celles de sa propre composition génétique”.
(Schéma inspiré de Vidyanond Nanjundiah, hénotic Plasticity, source ici).
Le mécanisme d’assimilation génétique selon Waddington: à gauche ce qu’on a l’impression qui se passe: un génotype G produisant un phénotype P se met à produire plein de nouveaux phénotypes suite à un changement d’environnement. La sélection d’un phénotype adapté (P’) se traduit par un changement dans le génotype (G’) après retour à l’environnement initial.L’explication (à droite): la population est composée de plusieurs génotypes (G1, G2...Gn) dont l’expression est canalisée pour produire un seul phénotype P. Cette canalisation est rompue suite à un changement d’environnement et de nombreux phénotypes sont produits. La sélection de P’ induit celle d’un nouveau sous-ensemble de génotypes dont le développement est canalisé pour produire P’.
Waddington ou Baldwin?
Si Waddington a bien été le premier à recréer expérimentalement ce phénomène d’assimilation génétique, d’autres avant lui avaient eu des idées semblables. James Baldwin en particulier, un psychologue américain du début du siècle, avait bien observé le “polyphénisme” de certaines espèces (le fait que leur morphologie peut changer en fonction de l’environnement). Mais alors que l’assimilation génétique de Waddington prédisait toujours une réduction de la variabilité des phénotypes, Baldwin était au contraire persuadé que l’évolution favorisait une plus grande plasticité dans le vivant. Il fallut attendre 2006 pour que le débat ne soit tranché par une très jolie expérience qui mit tout le monde d'accord.
Si Waddington a bien été le premier à recréer expérimentalement ce phénomène d’assimilation génétique, d’autres avant lui avaient eu des idées semblables. James Baldwin en particulier, un psychologue américain du début du siècle, avait bien observé le “polyphénisme” de certaines espèces (le fait que leur morphologie peut changer en fonction de l’environnement). Mais alors que l’assimilation génétique de Waddington prédisait toujours une réduction de la variabilité des phénotypes, Baldwin était au contraire persuadé que l’évolution favorisait une plus grande plasticité dans le vivant. Il fallut attendre 2006 pour que le débat ne soit tranché par une très jolie expérience qui mit tout le monde d'accord.
 L'histoire se passe cette fois avec les chenilles d'un sphynx vivant dans les plantes du tabac, Manduca sexta. Normalement Manduca est verte (photo de gauche en haut) mais il existe aussi une forme mutante, toute noire (photo du bas). Quand on chauffe ces chenilles pendant 6 heures à 42°, la variété verte reste verte, mais la variété mutante noire présente parfois des nuances de vert. Les chercheurs ont répété l’expérience sur plusieurs générations, en sélectionnant d’un côté les chenilles les plus polyphéniques (celles qui devenaient les plus vertes après le stress thermique), de l’autre les plus monophéniques (celles qui restaient immuables malgré le stress). Une troisième lignée sélectionnée au hasard sert de contrôle. Très vite on s'aperçoit que la lignée polyphénique change de couleur de plus en plus nettement (sur une échelle de 0 à 4, 4 étant la chenille entièrement verte et 0 entièrement noire) jusqu'à virer complètement au vert à la 13eme génération, alors que la lignée monophénique perd toute sensibilité au stress thermique dès la 7eme:
L'histoire se passe cette fois avec les chenilles d'un sphynx vivant dans les plantes du tabac, Manduca sexta. Normalement Manduca est verte (photo de gauche en haut) mais il existe aussi une forme mutante, toute noire (photo du bas). Quand on chauffe ces chenilles pendant 6 heures à 42°, la variété verte reste verte, mais la variété mutante noire présente parfois des nuances de vert. Les chercheurs ont répété l’expérience sur plusieurs générations, en sélectionnant d’un côté les chenilles les plus polyphéniques (celles qui devenaient les plus vertes après le stress thermique), de l’autre les plus monophéniques (celles qui restaient immuables malgré le stress). Une troisième lignée sélectionnée au hasard sert de contrôle. Très vite on s'aperçoit que la lignée polyphénique change de couleur de plus en plus nettement (sur une échelle de 0 à 4, 4 étant la chenille entièrement verte et 0 entièrement noire) jusqu'à virer complètement au vert à la 13eme génération, alors que la lignée monophénique perd toute sensibilité au stress thermique dès la 7eme:
On a ainsi réussi à créer artificiellement à la fois une lignée totalement monophénique de chenilles, insensibles aux chocs thermiques et une autre hyper polyphénique qui réagit brutalement à ces chocs. Les auteurs suggèrent que ce phénomène traduit le déplacement des niveaux de production d'hormone chez la chenille, par rapport à des seuils activant ou supprimant sa pigmentation (l'explication de PZ Myers dont j’ai tiré les illustrations, est limpide, si vous voulez creuser le sujet):
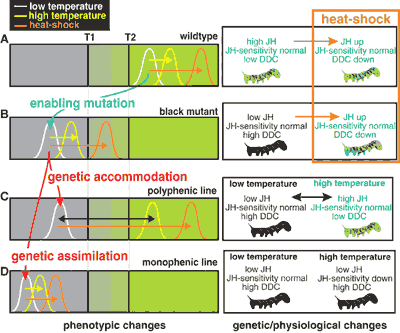
Cette expérience montre ainsi qu'assimilation génétique (lignée monophénique) et effet Baldwin (lignée polyphénique) ne sont que les facettes d'un même processus "d'accomodation génétique" grâce auquel le génotype laisse plus ou moins de flexibilité au développement de l'organisme. Ce qui a fait dire à certains biologistes que "le phénotype précède le génotype" dans l'évolution (même si il me semble qu’il a bien fallu une petite mutation au départ pour initier la machine).
Plasticité phénotypique + accommodation génétique = mécanisme évolutif
Sans nécessairement remettre en cause les grands principes de la théorie synthétique, ces feedbacks entre génétique et phénotype enrichissent considérablement l’idée qu’on se fait de l’évolution et constituent pour des biologistes comme Mary Jane West-Eberahrd, de puissants mécanismes de spéciations. Ainsi par exemple l’expérience sur les chenilles Maduca Sexta a-t-elle abouti à une lignée polyphénique (noire en dessous de 25°, verte au dessus de 30°) extrêmement semblable à une espèce voisine, Manduca quinquemaculata qui change de couleur exactement à ces températures!

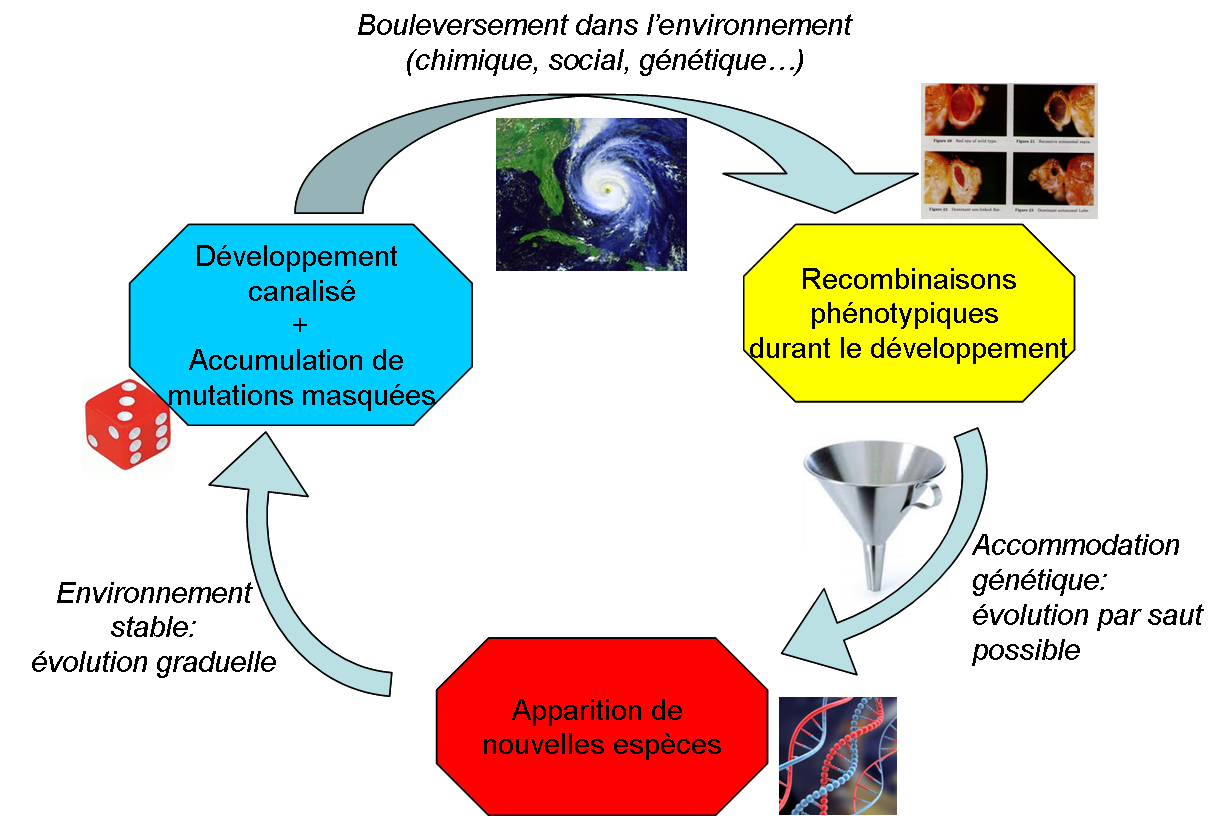
L’évolution vue par West-Eberhard
Une théorie séduisante à bien des titres
Ce mécanisme de “recombinaison phénotypique” suivie d’une “accomodation génétique” (vous excuserez mon jargon, mais j’espère que les concepts sont clairs) pourraient éclairer certaines zones d’ombre laissées par la théorie classique.
Par exemple, la rapidité avec laquelle s’observent certaines spéciations n’aurait rien de mystérieux: plus besoin d’attendre qu’un hasard salvateur produise une mutation favorable chez un individu, puis deux etc. La diversité génétique accumulée au sein d’une population permet en effet d’obtenir immédiatement un tas de phénotypes différents et les mêmes mutations favorables sont susceptibles d’apparaître chez plusieurs individus en même temps!
 |
| Les deux variétés d'épinoches Photo by Todd Hatfield (source here) |
La réapparition de certains traits ancestraux que l’on observe dans l’arbre du vivant s’expliquerait également assez bien si l’on considère ces traits non pas comme disparus mais comme “réprimés” à un stade donné de l’évolution. Il suffit là encore qu’un stress suffisamment fort réactive l’expression de ce trait pour le voir ressurgir tout au long de l’arbre de l’évolution.
Enfin la plus ou moins grande robustesse des mécanismes de canalisation expliquerait les immenses différences "d'évolutivité" entre des espèces à morphologie très changeante (les cychlidées dont je vous ai parlé dans mon billet précédent) et celles qui semblent immuables (les coelacanthes et les scorpions par exemple). Cette "absence de variabilité" est en réalité une très forte canalisation du développement en dépit des variations génétiques accumulées. Avez-vous remarqué que dans toutes les sciences, pour comprendre la stabilité des choses il faut souvent la considérer comme une situation d’équilbre entre des forces opposées (forces d'action et de réaction en physique, homéostasie en biologie, offre et demande en économie, etc?).
Sources:
Mary Jane West -berhard: Developmental plasticity and the origin of species differenciation (2005, pdf)
Mokzek & al, The role of developmental plasticity in evolutionary innovation (2011)
Crispo, The Baldwin effect and genetic assimilation (2007)
P Z Myers, Evolution of a polyphenism sur son blog Pharyngula
Billet connexe: l'épisode précédent, si vous l'avez loupé